Clarification
Our 2016 PNAS paper, “Life history effects on the molecular clock of autosomes and sex chromosomes”, has been the topic of an extensive discussion on Twitter over the past few days after Matthew Hahn, in a series of tweets, complained that our results are derivative of his, and that we failed to cite his earlier work on the topic, Thomas and Hahn 2014 MBE (T&H). We would like to set the public record straight about our results and their relationship to T&H’s.
We did not cite this paper because, due to an elementary arithmetical error, its derivation and main conclusion are fundamentally flawed[1], and we did not want to point people to a paper that is confusing and incorrect. T&H were informed of this error in 2014 but have chosen not to correct it in the record. Thus, readers following a direct citation from our paper would encounter their work without realizing that it is incorrect. However, a discussion of T&H and the correct derivation have been published elsewhere (Segurel, Wyman & Przeworski 2014 Annual Reviews of Human Genetics) and so we felt the correct choice for both scholarship and our readers was to cite T&H indirectly through Segurel et al. In retrospect, perhaps we should have also cited T&H and explained that the model is wrong and conclusions therefore unwarranted.
As part of his campaign against our work, Matthew Hahn posted a highly misleading figure (see below), implying more than a failure to cite them.
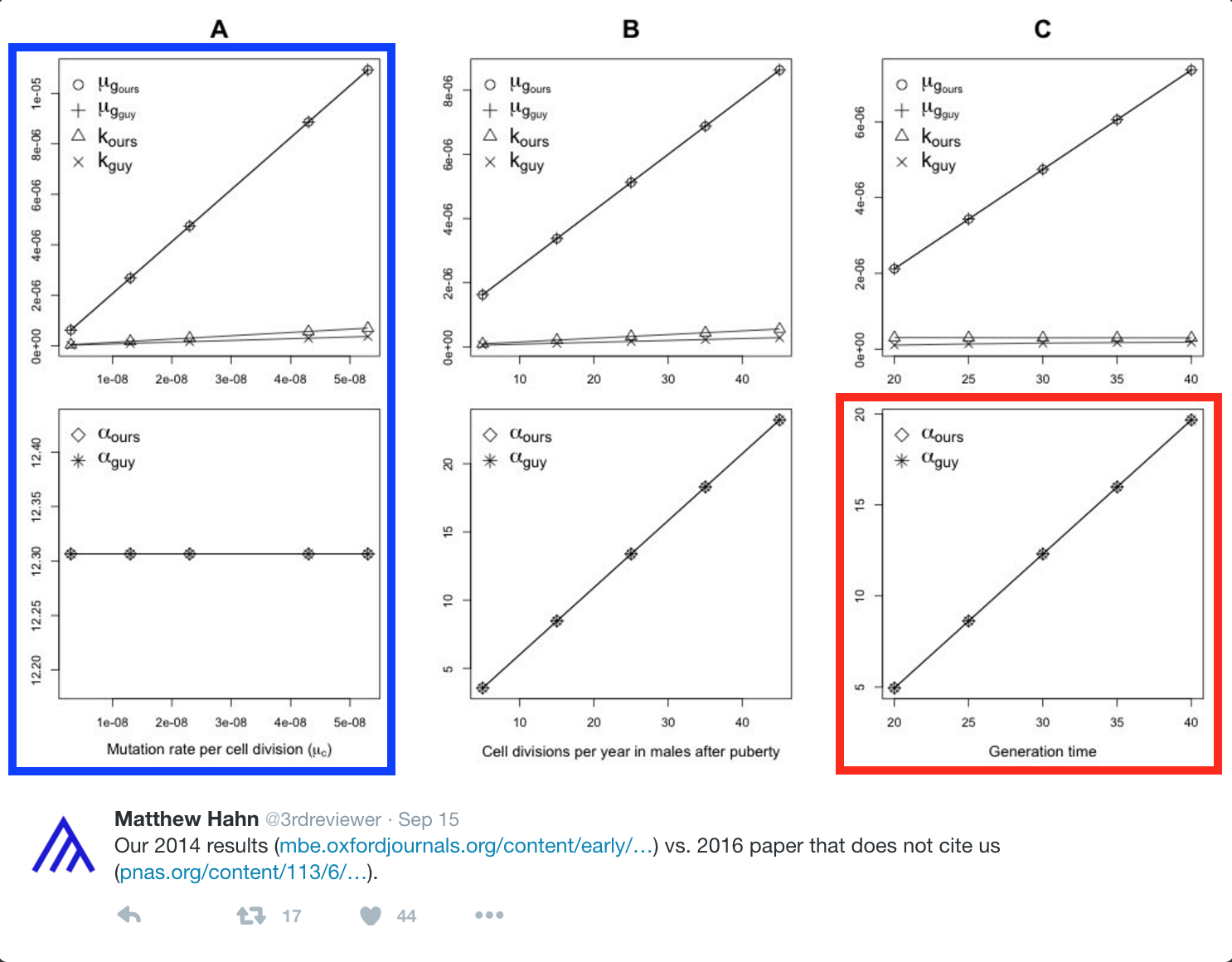
The figure and the parameter values he ascribes to us in the figure do not appear in our paper[2]. Moreover, the numbers that he places on the graph in our name (under “guy”) are incorrect summaries of our results[3] and reflect a complete misunderstanding of our paper[4].
Finally, beyond the general topic, there is very little overlap in our papers. The main points of our paper, as laid out in our abstract, are that (1) Sex differences in the generation time and other life history traits (e.g., the onset of puberty) have a major impact on the per year mutation rate, and that (2) Taking these factors into account can help to reconcile evolutionary dates based on mutations with our current understanding of the catarrhine fossil record. In contrast, T&H consider the same generation time in both sexes, do not consider the onset of puberty, and conclude (incorrectly) that life history traits should have negligible effects on the per year mutation rate. At no point do they relate their model to the fossil record.
If anyone is interested in more detail, we refer them to the two papers, as well as to Segurel et al. 2014 and other papers, such as Scally 2016 Phil. Trans Royal Society B.
Guy Amster & Guy Sella
[1] T & H focuses on how changing the generation time (i.e., the average age of reproduction) affects the yearly mutation rate. To this end, they set out to calculate the average number of de novo mutations in a child, as a function of the age of his or her parents at conception. To do so, they use the rates of cell divisions during three stages of the parents’ life history (pre-birth, post birth and post puberty) and a per cell division mutation rate. However, T&H’s calculation introduces an elementary arithmetic mistake: they calculate the yearly mutation rate in males as a simple average of yearly rates pre- and post-puberty (their supplemental equations 8-11), when the correct derivation weighs these rates by the time spent pre- and post- puberty (as laid out in Segurel et al. 2014, box 1). Critically, this arithmetic error leads them to erroneously conclude that the yearly mutation rate does not depend on the generation time (when in fact the generation time enters in the weighting of the rate post-puberty; again, see Segurel et al. 2014).
This mistake leads T&H to their main conclusion (see, e.g., their abstract): that the mutation rate per year should depend at most very weakly on the generation time (i.e., only through females and not at all through males, who contribute almost all the mutations under their assumptions). In the correct derivation, in contrast, a dependence of the mutation rate on generation time is expected under all but highly specific conditions on the underlying parameters and is therefore extremely likely (see box 2 of Segurel et al. 2014).
[2] As one example, we have highlighted in blue a plot that appears to examine the effect of changing the mutation rate per-cell division. We never vary this parameter in our paper. We are therefore at a loss to understand what Matthew Hahn is plotting in our name.
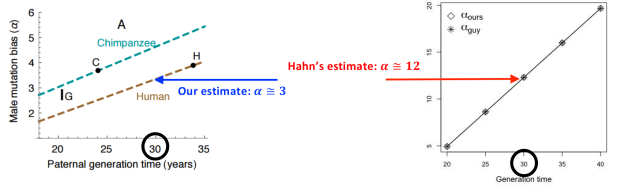
[3] This plot compares results from figure S4 in our paper (left) to what Matthew Hahn places on his graph in our name (right panel here; red box in the figure above).
[4] As one example, in the following tweet, Matthew Hahn states that α (the ratio of paternal to maternal mutations per generation) “is the same as” the X-to-autosome ratio of neutral divergence (by which we assume he means that there is a one to one correspondence, since they are obviously not equal):

In fact, one of the central findings of our paper is that once differences in generation time between males and females are taken into account, there is no longer any simple correspondence between alpha and the X to autosome ratio.
